( 五 ) 生物芯片
1991年Affymetrix公司的Fordor利用其所研發的光蝕刻技術制備了首個以玻片為載體的微陣列,標志著生物芯片正式成為可實際應用的分子生物學技術。時至今日,芯片技術已經得到了長足的發展,如果按結構對其進行分類,基本可分為基于微陣列( microarray) 的雜交芯片與基于微流控( microfluidic) 的反應芯片2 種。

1.微陣列芯片
( 1 ) 固相芯片: 微陣列基因組DNA 分析( microarray-based genomic DNAprofiling,MGDP) 芯片: 將微陣列技術應用于MGDP 檢測中已有超過十年的歷史,其技術平臺主要分為2 類,即微陣列比較基因組雜交( array-based comparative genomehybridization,aCGH) 和基因型雜交陣列( SNP array) 。顧名思義,aCGH 芯片使用待測DNA 與參比DNA的雙色比對來顯示兩者間的拷貝數變異( CNV) 的變化,而單核苷酸多態性( single nucleotide polymorphism,SNP) 芯片則無需與參比DNA 進行比較,直接通過雜交信號強度顯示待測DNA 中的SNP 信息。隨著技術的不斷進步,目前市場上已出現可同時檢測SNP 與CNV的高分辨率混合基因陣列芯片。MGDP 芯片主要應用于發育遲緩、先天性異常畸形等兒童遺傳病的輔助診斷及產前篩查。經驗證,使用MGDP 芯片進行染色體不平衡檢測與FISH 的診斷符合率可達100%,表達譜芯片( gene expression profiling array,GEP array) : 1999 年,Duggan 等首次使用cDNA 芯片繪制了mRNA 表達譜信息。隨著表觀遺傳學在疾病發生發展中的作用日益得到重視,目前也已出現microRNA 芯片、長鏈非編碼RNA( long noncoding RNA,lncRNA) 芯片等。類似于MGDP 芯片,GEP 芯片使用反轉錄后生成的cDNA 文庫與固定于芯片載體上的核酸探針進行雜交,從而檢測雜交熒光信號的強度判斷基因的表達情況。相較于基因組雜交,GEP 芯片對生物學意義更為重要的轉錄組信息進行檢測,對疾病的診斷與預后判斷具有特殊的意義。目前使用GEP 芯片對急性髓細胞白血病、骨髓增生異常綜合征等血液病及神經退行性變等進行診斷、分類及預后評估已經獲得了令人滿意的效果;
( 2 ) 液相芯片:傳統固相芯片將檢測探針錨定于固相載體上捕獲目的序列,而Luminex 公司的xMAP 技術則通過搭配不同比例的2 種紅色熒光染料,將聚苯乙烯微球標記為不同的熒光色,并對其進行編碼得到具有上百種熒光編號的微球。通過xTAG 技術將不同的特異性雜交探針交聯至編碼微球上,使得不同的探針能夠通過微球編碼得以區分。利用混合后的探針-微球復合物與待測樣本進行雜交,使微球在流動鞘液的帶動下通過紅綠雙色流式細胞儀,其中紅色激光檢測微球編碼,綠色熒光檢測經雜交后核酸探針上熒光報告基團的信號強度,一次完成對單個樣本中多種靶序列的同時鑒定。目前,該技術已在囊性纖維化等遺傳性疾病診斷、多種呼吸道病毒鑒定及人乳頭瘤病毒分型取得了廣泛的應用。
2.微流控芯片
1992 年Harrison 等首次提出了將毛細管電泳與進樣設備整合到固相玻璃載體上構建“微全分析系統”的構想,通過分析設備的微型化與集成化,完成傳統分析實驗室向芯片上實驗室( lab-on-chip) 的轉變。微流控芯片( microfluidic chip) 由微米級流體的管道、反應器等元件構成,與宏觀尺寸的分析裝置相比,其結構極大地增加了流體環境的面積/體積比,以最大限度利用液體與物體表面有關的包括層流效應、毛細效應、快速熱傳導和擴散效應在內的特殊性能,從而在1 張芯片上完成樣品進樣、預處理、分子生物學反應、檢測等系列實驗過程。
目前使用微流控芯片進行指導用藥的多基因位點平行檢測是主要臨床應用領域。
二、核酸序列測定
測序反應是直接獲得核酸序列信息的唯一技術手段,是分子診斷技術的一項重要分支。雖然分子雜交、分子構象變異或定量PCR 技術在近幾年已得到了長足的發展,但其對于核酸的鑒定都僅僅停留在間接推斷的假設上,因此對基于特定基因序列檢測的分子診斷,核酸測序仍是技術上的金標準。
( 一 ) 第1代測序
1975年Sanger與Coulson 發表了使用加減法進行DNA序列測定的方法,隨后Maxam在1977年提出了化學修飾降解法的模型,為核酸測序時代的來臨拉開了序幕。
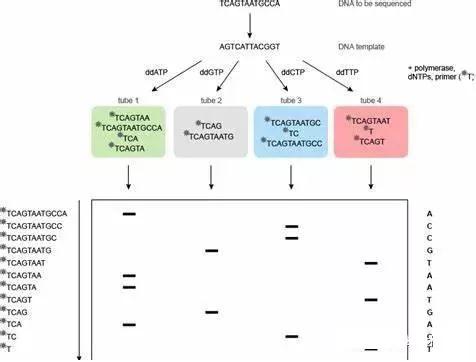
Sanger 等于同年提出的末端終止法( Sanger測序法) 利用2'與3'不含羥基的雙脫氧核苷三磷酸( ddNTP) 進行測序引物延伸反應,ddNTP在DNA 合成反應中不能形成磷酸二酯鍵,DNA合成反應便會終止。如果分別在4 個獨立的DNA 合成反應體系中加入經核素標記的特定ddNTP,則可在合成反應后對產物進行聚丙烯酰胺凝膠電泳( polyacrylamide gelelectrophoresis,PAGE) 及放射自顯影,根據電泳條帶確定待測分子的核苷酸序列。Appied Biosystems 公司在Sanger法的基礎上,于1986 年推出了首臺商業化DNA 測序儀PRISM 370A,并以熒光信號接收和計算機信號分析代替了核素標記和放射自顯影檢測體系。該公司于1995 年推出的首臺毛細管電泳測序儀PRISM 310 更是使測序的通量大大提高。Sanger 測序是最為經典的一代測序技術,仍是目前獲取核酸序列最為常用的方法。
( 二 ) 第2代測序
1.焦磷酸測序(Pyro-sequencing)
不同于Sanger 測序法所使用的合成后測序理念,Ronaghi分別于1996 年與1998 年提出了在固相與液相載體中通過邊合成邊測序的方法-焦磷酸測序。其基本原理是利用引物鏈延伸時所釋放的焦磷酸基團激發熒光,通過峰值高低判斷與其相匹配的堿基數量。由于使用了實時熒光監測的概念,焦磷酸測序實現了對特定位點堿基負荷比例的定量,因此在SNP 位點檢測、等位基因( 突變)頻率測定、細菌和病毒分型檢測方面應用廣泛。由于熒光報告原理不同,其對于序列變異的檢測靈敏度從Sanger 測序的20%提高到了5%。但由于該技術的儀器采購與單次檢測成本較高,目前尚未得到大規模的臨床使用。
2.高通量第2代測序
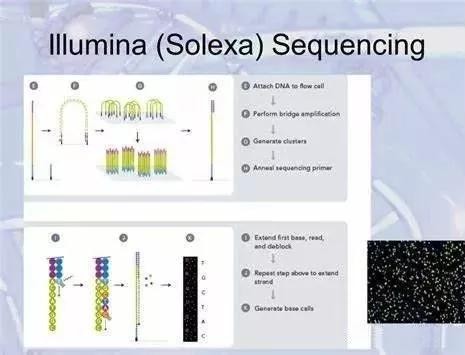
目前常見的高通量第二代測序平臺主要有Roche 454、Illumina Solexa、ABI SOLiD 和Life Ion Torrent 等,其均為通過DNA 片段化構建DNA 文庫、文庫與載體交聯進行擴增、在載體面上進行邊合成邊測序反應,使得第1 代測序中最高基于96 孔板的平行通量擴大至載體上百萬級的平行反應,完成對海量數據的高通量檢測。該技術可以對基因組、轉錄組等進行真正的組學檢測,在指導疾病分子靶向治療、繪制藥物基因組圖譜指導個體化用藥、感染性疾病的病原微生物宏基因組鑒定及通過母體中胎兒DNA 信息進行產前診斷等方面已經取得了喜人的成績。然而,由于該技術需要對DNA 進行片段化處理,測序反應讀長較短( 如Solexa 與SOLiD系統單次讀長僅50bp) ,需要對數據進行大規模拼接,因此對分子診斷工作者掌握生物信息學知識提出了更高要求,以利于后期的測序數據分析。
( 三 ) 第3代測序
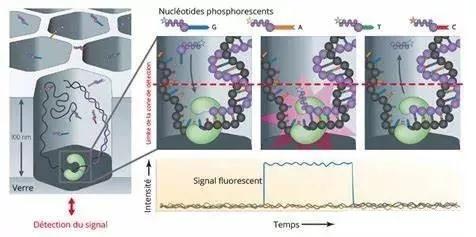
第3 代測序技術的核心理念是以單分子為目標的邊合成邊測序。該技術的操作平臺目前主要有Helicos 公司的Heliscope、Pacific Biosciences 公司的SMRT 和Oxford Nanopore Technologies 公司的納米孔技術等。該技術進一步降低了成本,可對混雜的基因物質進行單分子檢測,故對SNP、CNV 的鑒定更具功效。但是目前其進入產品商業化,并最終投入臨床應用仍有很長的距離。
三、基于分子構象的分子診斷技術
( 一 ) 變性梯度凝膠電泳( denaturing gradient gel electrophoresis,DGGE )與單鏈構象多態性( single strand conformationpolymorphism,SSCP)
1970 ~1980 年間,Fischer 等與Orita 等分別提出了利用核酸序列變異所導致雙鏈變性條件差異與單鏈空間折疊差異,通過變性與非變性PAGE 對變異序列進行分離鑒定的方法,即DGGE與SSCP。上述2 項技術均通過變異核酸分子在空間構象上的差異,通過特定條件下電泳速率的變化進行檢測。正因為核酸分子構象具有序列特異性,且對于序列的改變非常敏感,常常1 個堿基的變化也能得到鑒定。但由于DGGE 與SSCP 均必須進行PCR 后開蓋電泳的操作,現已不常見于臨床檢測。
(二) 變性高效液相色譜 (denaturing high-performance liquid chromatography,dHPLC)
1997 年,Oefner 和Underhill 建立了利用異源雙鏈變性分離變異序列、使用色譜洗脫鑒定的技術,稱為dHPLC,可自動檢測單堿基置換及小片段核苷酸的插入或缺失。對于存在一定比例變異序列的核酸雙鏈混合物,其經過變性和復性過程后,體系內將出現2 種雙鏈: 一種為同源雙鏈,由野生正義鏈-野生反義鏈或變異正義鏈-變異反義鏈構成的核酸雙鏈; 另一種為異源雙鏈,即雙鏈中1 條單鏈為野生型,而另1 條為變異型。由于存在部分堿基錯配的異源雙鏈DNA 與同源雙鏈DNA 的解鏈特征不同,在相同的部分變性條件下,異源雙鏈因存在錯配區而更易變性,被色譜柱保留的時間短于同源雙鏈,故先被洗脫下來,從而在色譜圖中表現為雙峰或多峰的洗脫曲線。由于該技術使用了較高分析靈敏度的色譜技術進行檢測,可快速檢出< 5%負荷的變異序列。但需注意的是,由于該技術主要通過異源雙鏈進行序列變異檢測,其不能明顯區分野生型與變異型的純合子。
(三)高分辨率熔解分析( high-resolution melting analysis,HRMA)
2003年,Wittwer 等首次革命性地使用過飽和熒光染料將PCR 產物全長進行熒光被動標記,再通過簡單的產物熔解分析對單個堿基變化進行鑒定。該技術的原理也是通過異源雙鏈進行序列變異鑒定。待測樣本經PCR 擴增后,若存在序列變異雜合子,則形成異源雙鏈,其熔解溫度大大下降。此時由于雙鏈被飽和染料完全填充,其產物熔解溫度的變化便可通過熔解曲線的差異得以判定。對于變異純合子而言,HRMA 也可利用其較高的分辨率完成PCR 產物單個位點A: T 雙鍵配對與G: C 三建配對熱穩定性差異的鑒定,但是對于Ⅱ、Ⅲ類SNP 的純合子變異則無法有效區分。
如何利用DNA 構象對序列進行推測,從而避免成本較高的序列測定或操作繁瑣的雜交反應一直是分子生物學研究與應用的熱點問題。目前,使用構象變化對序列變異進行間接檢測的便捷性已得到一致肯定,尤其是HRMA 可完成對變異序列單次閉管的擴增檢測反應。但需要注意的是,由于基于構象變化的分子檢測手段多無法通過探針雜交或核酸序列測定對檢測的特異性進行嚴格的保證,因此其只適合大規模的初篩,而真正的確診仍需要進行雜交或測序的驗證。
四、定量PCR( quantitative PCR,qPCR)
相比于其他分子診斷檢測技術,qPCR 具有2項優勢,即核酸擴增和檢測在同一個封閉體系中通過熒光信號進行,杜絕了PCR 后開蓋處理所帶來擴增產物的污染; 同時通過動態監測熒光信號,可對低拷貝模板進行定量。正是由于上述技術優勢,qPCR 已經成為目前臨床基因擴增實驗室接受程度最高的技術,在各類病毒、細菌等病原微生物的鑒定和基因定量檢測、基因多態性分型、基因突變篩查、基因表達水平監控等多種臨床實踐中得到大量應用。但伴隨著qPCR 技術的迅猛發展,有關這項技術的質量管理問題也日益突出,如何消除各類生物學變量所引起的檢測變異,減少或抑制實驗操作與方法學中的各種干擾因素是qPCR 技術面臨的難題。
( 一 ) 實時熒光定量PCR(real-time PCR)
1.雙鏈摻入法
1992 年Higuchi 等通過在PCR 反應液中摻入溴乙錠對每個核酸擴增熱循環后的熒光強度進行測定,提出了使用熒光強度與熱循環數所繪制的核酸擴增曲線,定量反應體系中初始模板的反應動力學( real-time PCR)模型,開創了通過實時閉管檢測熒光信號進行核酸定量的方法。核酸染料可以嵌入DNA 雙鏈,且只有嵌入雙鏈時才釋放熒光,在每1 次的擴增循環后檢測反應管的熒光強度,繪制熒光強度-熱循環數的S 形核酸擴增曲線,以熒光閾值與擴增曲線的交點在擴增循環數軸上的投影作為循環閾值( Cycle threshold,Ct) ,則Ct 與反應體系中所含初始模板數量呈負指數關系,推斷初始模板量。隨后Morrison提出了使用高靈敏度的雙鏈染料SYBR Green I 進行反應體系中低拷貝模板定量的方法。這一方法操作簡便,但由于僅使用擴增引物的序列啟動核酸擴增,其產物特異性無法得到充分保證。雖然在實時熒光定量PCR 反應后可通過熔解曲線對產物特異性進行檢驗,但其特異性明顯遜于使用熒光探針進行檢測,因此雙鏈摻入法并未在臨床實踐中得到認可。
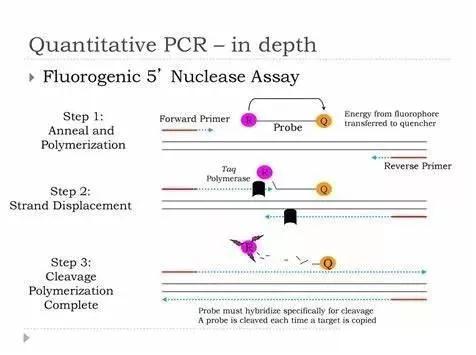
2. Taqman 探針
由于雙鏈摻入法存在特異性較低的問題,1996 年Heid綜合之前發現的Taq 酶的5'核酸酶活性與熒光共振能量轉移( fluorescence resonance energytransfer,FRET) 探針的概念提出了使用Taqman 探針進行qPCR 的方法。TaqMan 探針的本質是FRET 寡核苷酸探針,在探針的5'端標記熒光報告基團,3'端標記熒光淬滅基團,利用Taq 酶具有5' ~ 3'外切酶活性,在PCR過程中水解與靶序列結合的寡核苷酸探針,使熒光基團得以游離,釋放熒光信號。從而使能夠與靶序列雜交的探針在擴增過程中釋放熒光,通過real-time PCR 的原理對其進行定量。由于其超高的特異性與成功的商品化推廣,Taqman 探針已經成為目前臨床使用最為廣泛的qPCR 方法,其在各種病毒基因定量檢測、基因分型、腫瘤相關基因表達檢測等方面具有著不可替代的地位。
3.分子信標
同樣在1996 年,Tyagi 等提出了使用分子信標( moleuclar beacons) 進行qPCR的方法,分子信標是5'與3'端分別標記有熒光報告基團與淬滅基團的寡核苷酸探針,其兩端具有互補的高GC 序列,在qPCR 反應液中呈發夾結構,熒光基團與淬滅基團發生熒光共振能量轉移( FRET) 而保持靜息狀態。當PCR 反應開始后,莖環結構在變性高溫條件下打開,釋放熒光;在退火過程中,靶序列特異性探針則與模板雜交保持線性,不能與模板雜交的探針則復性為莖環結構而熒光淬滅,通過檢測qPCR 體系中退火時的熒光信號強度,便可以real-time PCR原理特異性檢測體系中的初始模板濃度。相比于Taqman 探針,分子信標使用發卡結構使熒光基團與淬滅基團在空間上緊密結合,大大降低了檢測的熒光背景,其檢測特異性較Taqman 探針更高,更適合等位基因的分型檢測。
4.雙雜交探針
1997 年,Wittwer 等發表了使用分別標記熒光供體基團與熒光受體基團的2 條相鄰寡核苷酸探針進行qPCR 的方法。雙雜交探針所標記的供體基團和受體基團的激發光譜間具有一定重疊,且2 條探針與靶核酸的雜交位置應相互鄰近。僅當2 條探針與靶基因同時雜交時,供體與受體基因得以接近,從而通過FRET 發生能量傳遞,激發熒光信號,熒光信號強度與反應體系中靶序列DNA 含量呈正比。由于使用了2條探針進行靶序列雜交,該方法的特異性比傳統單探針檢測體系得到了極大地提升。
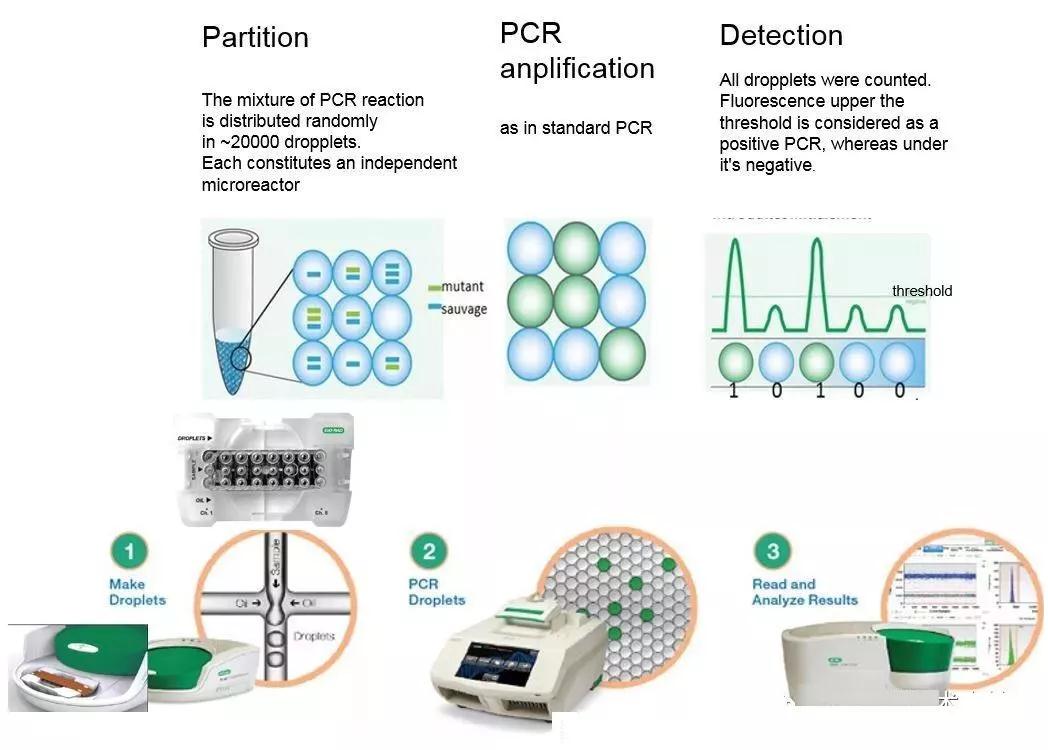
( 二 ) 數字PCR
早在上世紀90 年代就出現了使用微流控陣列對單次qPCR 反應進行分散檢測的概念。基于這一理念,Vgelstein 與Kinzter于1999 年發表了數字PCR( digital PCR) 的方法,對結腸癌患者糞便中的微量K-RAS 基因突變進行了定量。相比于傳統的qPCR 方法,數字PCR 的核心是將qPCR 反應進行微球乳糜液化,再將乳糜液分散至芯片的微反應孔中,保證每個反應孔中僅存在≤1 個核酸模板。經過PCR 后,對每個微反應孔的熒光信號進行檢測,存在靶核酸模板的反應孔會釋放熒光信號,沒有靶模板的反應孔就沒有熒光信號,以此推算出原始溶液中待測核酸的濃度。因此,數字PCR 是1 種檢測反應終點熒光信號進行絕對定量的qPCR反應,而非以模板Ct 值進行核酸定量的real-time PCR。
經由Quantalife 公司開發( 已于2011 年被BIO-RAD 收購) 的微滴式數字PCR 是首款商品化的數字PCR 檢測系統,目前已被廣泛運用于微量病原微生物基因檢測、低負荷遺傳序列鑒定、基因拷貝數變異與單細胞基因表達檢測等多個臨床前沿領域。與傳統qPCR 相比較,該技術具有超高的靈敏度與精密度,使其成為目前qPCR 領域的新星。
半個世紀以來分子診斷的高速發展離不開分子生物學技術日新月異的進步。概而言之,在過去的50 年中分子診斷技術取得了三大轉化與3項提升: 即報告信號檢測從放射核素標記向熒光標記轉化,操作方法由手工操作向全自動化轉化,檢測分析通量從單一標志物向高通量多組學聯合判斷轉化; 檢測靈敏度、精密度、特異性的快速提升。
在未來5年中,我國分子診斷事業將迎來兩方面的進步。隨著衛生監管部門對分子診斷重要性的認識不斷深入與越來越多高學歷、高素質人才的進入,分子診斷將會出現理念的革命性進步,高通量技術將更多的進入臨床的實際應用中。隨著技術的進一步發展,傳統針對特定基因異常、病原微生物感染鑒定的方法學,也將在檢測的各項分析性能與操作便捷程度上取得長足的進步。對于傳統人力與時間成本較高的檢測方法學,將出現兩極分化的態勢,即Southern 等經典的檢測金標準將得到保留; 而ASO-RDB 等靈敏度、特異性均不能滿足實際臨床需求的方法將快速被新型技術所取代。最終,分子診斷也必將一改目前僅僅用于病原微生物基因檢測與部分遺傳性疾病診斷的局面,形成由腫瘤學、遺傳學、微生物學、藥物基因組學四足鼎立,快速發展的景象。